Engineered Virus-Like Particles Redefine...
CMN Special
Feb. 25, 2026


CMN Intelligence - The World’s Most Comprehensive Intelligence Platform for CRISPR-Genomic Medicine and Gene-Editing Clinical Development
Providing market intelligence, data infrastructure, analytics, and reporting services for the global gene-editing sector. Read more...
Prime editing was described initially by the laboratory of Dr. David Liu in 2019 (1). This powerful tool combines the DNA-scanning and sequence-identification capabilities of the CRISPR-Cas9 system with a reverse transcriptase enzyme, which uses an RNA template to synthesise a new single-strand DNA (ssDNA) sequence and insert it into the DNA.
Prime editors (PEs) can introduce any of the twelve possible types of nucleotide substitutions using the same PE protein configuration (1). This allows greater flexibility than base editing, which requires a different protein configuration for each of the four types of nucleotide substitutions that are feasible to date (see our explainer about base editing for more details).
In addition, PEs can also introduce small-to-medium-sized targeted insertions and deletions with no or very few unwanted modifications at the target site, and virtually no off-target effects (2). This is because PEs, unlike other CRISPR-Cas9-mediated targeted integration approaches, do not generate DSBs as part of their gene-editing process. This greatly reduces the potential for DSB-related chromosomal aberrations and cell toxicity, which are two important limiting factors for safe gene editing.
Similarly to base editors, PEs are generated by coupling two separate proteins capable of very specific functions, as illustrated in Figure 1. The components of a typical prime editor include:
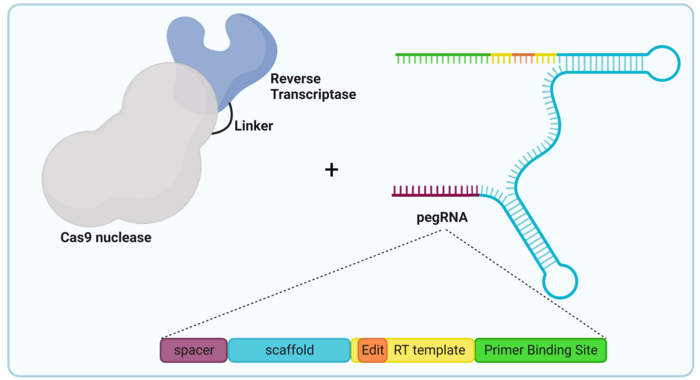
Prime editing is a complex process that involves a nick of the non-complementary strand and the hybridisation of the pegRNA and the nicked strand, which serves as a primer for reverse transcription (Ref. 2, see Figure 2).
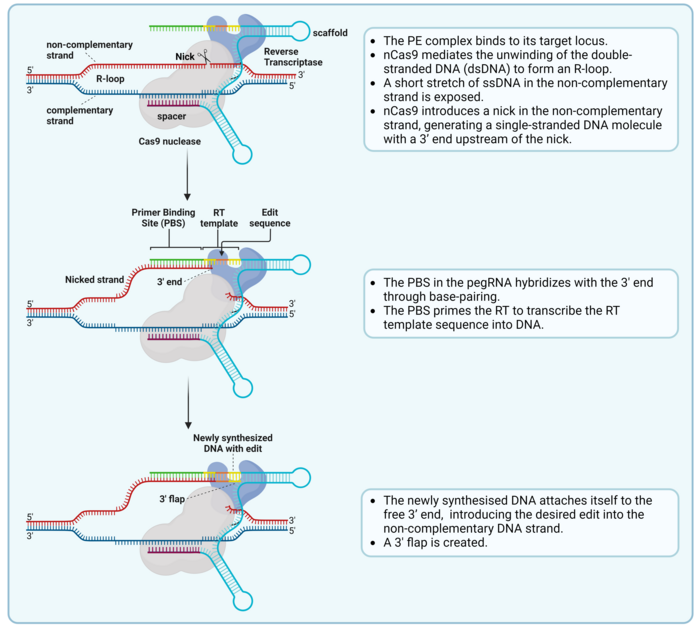
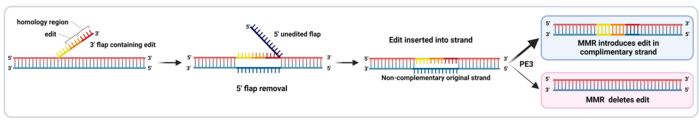
The final part of the RT template sequence includes a region of homology with the DNA sequence downstream of the nick, which allows the 3’ flap to align with the complementary strand (Ref 2, see Figure 3). To make room for this alignment, the 5’ end that was present downstream of the nick in the target sequence is detached from the complementary strand as a 5’ flap and removed by endogenous cellular DNA repair mechanisms. Then, the remaining nick is repaired, and a new double-stranded DNA (dsDNA) molecule is formed (Figure 3).
However, since the introduced edit is only present in the non-complementary strand, the new dsDNA contains two strands with different information in them. This causes incomplete alignment between both strands, also known as a mismatch. This is repaired by the mismatch repair (MMR) machinery, which randomly takes one of the two strands as a template and repairs the other strand to match it. This means that in half the cases, the MMR will reject the edit and re-construct the original sequence. However, a newer PE version called PE3 includes an extra sgRNA, which is used by the PE complex to nick the strand which does not include the edit. This leads to the MMR machinery preferentially using the edited strand as a repair template, which introduces the edit in the other strand and generates the final dsDNA edited sequence (Figure 3). PE3 introduces edits with a 4.2-fold higher efficiency than PE2 (1,2).
Several fascinating therapeutic approaches based on prime editing have been explored all over the world in the last few years. For example, in August 2021, the group of Hyongbum Henry Kim (Yonsei University) published the first report of successful in vivo prime editing both in the retina and in the liver (3), achieving correction of mutations and phenotypes in mouse models of Leber’s congenital amaurosis and hereditary tyrosinemia, respectively. Similarly, the group of Gerald Schwank (University of Zurich) reported in March 2022 that they performed liver-directed prime editing in a mouse model of phenylketonuria, achieving therapeutic reduction of blood phenylalanine (4). The group of David Liu (Harvard University) has also recently reported successful prime editing in the mouse brain, liver and heart (5).
Beyond therapeutic applications, scientists are also exploring the possibility of using prime editing for other applications, including the generation of disease models, screening assays, and very importantly, the generation of crops with desired properties that can facilitate agriculture in the future.
Despite its advantages over other gene-editing technologies, prime editing presents some limitations that are currently being addressed by scientists all over the world (2,6).
Some of the limitations are similar to those of conventional Cas9 editing approaches regarding targeting and delivery. Thus, already existing Cas9 variants that were developed to solve those issues can be used to construct prime editors able to overcome the same issues. These new PE variants can expand the range of target DNA sequences that can be edited, but they are generally less efficient than the original PEs, which suggests that further optimisation is required.
When it comes to in vivo delivery, the increased size of the PE complex compared to regular Cas9 complicates its delivery. To address this hurdle, the intein system, which is used to package other Cas9 products into adeno-associated viral vectors, has also been used for in vivo delivery of base editors. Other authors have also used ribonucleoprotein complexes embedded in virus-like particles, which overcomes packaging issues and guarantees short-term expression of the PE (7). However, due to the nature of prime editing, its other limitations are quite different from those of other gene editing approaches (2):
Stay tuned for future explainers about various gene-editing technologies. In the meantime, you can find all of our coverage on prime editing right here.
1. Anzalone, A. V. et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature. 2019. 576(149–157).
2. Chen, P. J. & Liu, D. R. Prime editing for precise and highly versatile genome manipulation. Nat. Rev. Genet. 2023. 24(161–177).
3. Jang, H. et al. Application of prime editing to the correction of mutations and phenotypes in adult mice with liver and eye diseases. Nat. Biomed. Eng. 2022. 6(181–194).
4. Böck, D. et al. In vivo prime editing of a metabolic liver disease in mice. Sci. Transl. Med. 2022. 14(eabl9238).
5. Davis, J. R. et al. Efficient prime editing in mouse brain, liver and heart with dual AAVs. Nat. Biotechnol. 2023. doi:10.1038/s41587-023-01758-z.
6. Zhao, Z. et al. Prime editing: advances and therapeutic applications. Trends Biotechnol. 2023. 41(1000–1012).
7. Haldrup, J. et al. Engineered lentivirus-derived nanoparticles (LVNPs) for delivery of CRISPR/Cas ribonucleoprotein complexes supporting base editing, prime editing and in vivo gene modification. Nucleic Acids Res. 2023. 51(18):10059–10074.
Manel Llado is a gene editing expert and science writer/communicator based in Vienna, Austria
We acknowledge Dr. Jakob H. Haldrup (Radcliffe Department of Medicine, Oxford) for advice and input.